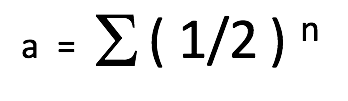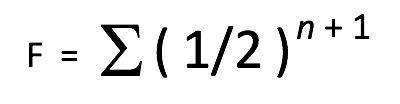Pedigrees
 Pedigrees are the official record of the parentage and thus the ancestry of an animal. They are an integral part of the official record keeping required by a breed club to register new animals. Pedigrees are also a valuable resource for the genetics of an animal, whereby the genetic contribution of a particular ancestor can be estimated and the genetic relatedness of two individuals can be determined. As we will see, the pedigree is also used for determining inbreeding estimates for animals.
Pedigrees are the official record of the parentage and thus the ancestry of an animal. They are an integral part of the official record keeping required by a breed club to register new animals. Pedigrees are also a valuable resource for the genetics of an animal, whereby the genetic contribution of a particular ancestor can be estimated and the genetic relatedness of two individuals can be determined. As we will see, the pedigree is also used for determining inbreeding estimates for animals.
Pedigrees are organized like the branches of a tree. In the standard format, the paternal contribution is presented above and the maternal contribution is presented below for the animal (offspring) in question. With each generation, the genetic contribution of a particular ancestor is divided by two. Thus, an animal (offspring) has 2 parents each of whom contributed 50% to the genetic constitution of their offspring and 4 grandparents each of whom contributed 25% to the genetic constitution of the offspring, and so on.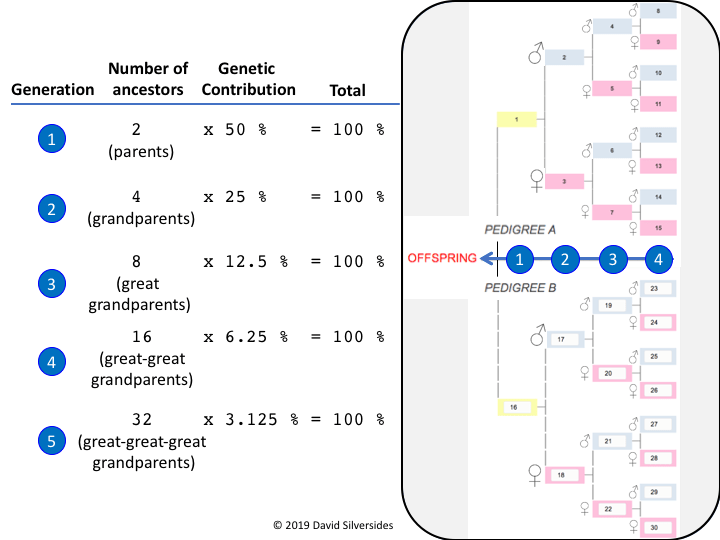
Inbreeding
When two animals that share common ancestors are bred together, a condition of inbreeding exists in the offspring. This inbreeding has two consequences for the phenotype of the offspring:
- Increased uniformity of “type”(i.e. phenotype) within the offspring, with increased “prepotency”, i.e. the ability of parents to transmit or fix a phenotype in the next generation. This uniformity of type is desirable to breeders.
- Inbreeding depression, which includes a reduction of vitality, reduced weight, reduced fertility, reduced rate of growth; increase rates of congenital anomalies, increased mortality, increased rates of recessive genetic diseases; a shortened life span. Inbreeding depressionis cumulative: with increased inbreeding and over time there is an increase in inbreeding depression.
 At the level of the genome, inbreeding has the effect of increasing the percentage of homozygous genetic alleles (N/N, M/M) and reducing the percentage of heterozygous genetic alleles (M/N, carriers). Mutations that are recessive will accumulate in the homozygous state (M/M), thus increasing the frequency of recessive genetic disease, which contributes to the deleterious effects of inbreeding depression. See Horse Genetics 3.0 Simple Genetic Diseases.
At the level of the genome, inbreeding has the effect of increasing the percentage of homozygous genetic alleles (N/N, M/M) and reducing the percentage of heterozygous genetic alleles (M/N, carriers). Mutations that are recessive will accumulate in the homozygous state (M/M), thus increasing the frequency of recessive genetic disease, which contributes to the deleterious effects of inbreeding depression. See Horse Genetics 3.0 Simple Genetic Diseases.
Stated simply, Mother Nature does not like uniformity, neither in phenotypes nor in genotypes. She much prefers variations, diversity and differences, for these are the motors of evolution.
A limited number of foundation animals, closed pedigree books for a purebred breed and the tendency to use only a fraction of animals within a breed as parents for the next generation (popular sire effect) all contribute to the reality that purebred horse breeds often represent a more or less inbred population of animals.
The Coefficient of Relation (a) and the Coefficient of Inbreeding (COI)
In 1921-22, Sewall Wright, an American mathematician and geneticist, defined two mathematical values based on a given genetic pedigree: the Coefficient of Relation (a) and the Coefficient of Inbreeding (COI, inbreeding coefficient or sometimes simply “F”). These two values are useful for the art and science of domestic animal breeding. Neither of these values represent specific DNA or genes of an animal. They are abstract values, calculated on the assumption that all of our genes follow simple mendelian genetics with dominant and recessive alleles (versions). This assumption is of course simplistic. In spite of this, the Coefficient of Relation and the Coefficient of Inbreeding have proven to be useful measures with practical applications for animal and plant breeders. They allow an estimation of the genetic relationship between two animals and also an estimation of the genetic variation (or lack thereof) found within the genome of a particular animal. With these measures, breeders can estimate the risks of having undesirable health effects (inbreeding depression) in future offspring and generations due to a lack of genetic variation caused by too much inbreeding.
The Coefficient of Relation (a) is an estimation of the quantity of DNA that is in common between two animals within a pedigree. The simplest formula representing the coefficient of relation is as follows:
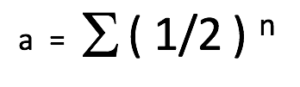
Where:
a = the Coefficient of Relation between two animals.
½ = the genetic contribution of a parent towards its offspring.
n = the number of pathways (number of generations) that separate two animals within a pedigree. Stated differently, n is the number of meiosis (cell division of sex cells) separating two animals. In other words, the number of times productive sex occurred between the ancestors (or descendants) of the two animals in question.
∑ = the sum of the calculations for all possible pathways linking the two individuals.
This equation is derived from the fact that we (and our horses) are diploid, and that we have received half of our DNA from our mother and the other half from our father. The DNA of our parents was divided in two (1/2) within their germ cells, and then added together (1/2 + 1/2) during fertilization to generate the full double (diploid) genetic complement (100% or 1.0) needed to form us and make us function. If two animals are not genetically linked then their Coefficient of Relation is 0. On the other hand, if two animals are linked within a pedigree, their Coefficient of Relation will be higher than 0 and can be calculated.
The Coefficient of Inbreeding (COI, inbreeding coefficient, “F”) is an estimation of the loss of genetic variation for an individual animal, due to the fact of having a common ancestor on both the paternal side and the maternal side of its pedigree. The Coefficient of Inbreeding can be derived from the Coefficient of Relation by the following simple formula:
COI = (1/2) a = (1/2) n+1
A more comprehensive presentation of the Coefficient of Inbreeding formula is as follows:
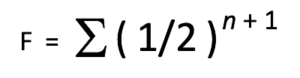
Where:
F = the Coefficient of Inbreeding (COI) for the individual in question.
½ = the genetic contribution of a parent towards its offspring.
n = the number of pathways (number of generations) between a common ancestor and the individual in question.
+1 = an additional factor of (1/2) is added to represent the anticipated loss of genetic diversity due to common ancestors on both maternal and paternal sides of the pedigree.
∑ = the sum of the calculations for each individual ancestor in common.
If the common ancestor has itself common ancestors, then the common ancestor will itself have a positive inbreeding coefficient value. The inbreeding coefficient for the animal in question is now multiplied by the following correction factor:
(1 + Fa)
Where:
Fa = the Coefficient of Inbreeding for the ancestor in common.
For intrepid souls, mathematical masochists, mad dogs and Englishmen, these formulas can get even more complex:
http://www.genetic-genealogy.co.uk/Toc115570144.html
http://www.genetic-genealogy.co.uk/Toc115570148.html.
Examples of Coefficients of Relation and Coefficient of Inbreeding
Here are some examples of Coefficients of Relation (once again, the average amount of shared DNA) between two parents without common ancestors, as well as the Coefficient of Inbreeding (measurement of loss of genetic diversity) for their hypothetical offspring:
| Relationship |
Coefficient of Relation (a) |
Coefficient of Inbreeding (COI)
of offspring |
| Parent – child |
50% |
25% |
| Brother – sister |
50% |
25% |
| Grandparent – grandchild |
25% |
12.5% |
| Uncle/aunt – nephew/niece |
25% |
12.5% |
| Half brother – half sister |
25% |
12.5% |
| Cousins |
12.5% |
6.25% |
| Half-cousins |
6.25% |
3.125% |
| Second cousins |
3.13% |
1.063% |
For example, a child shares 50% of its genes with its parent, and the Coefficient of Relation (a) between the parent and child is 50%. The child has a Coefficient of Inbreeding of 0. However, the offspring resulting from an incestuous relationship between a parent and their child, or between a brother and sister, will have a Coefficient of Inbreeding of 25%. This means that, on average, there will be a 25% loss of genetic variation (loss of M/N) and an equivalent 25% gain in genetic uniformity (gain in N/N, M/M) at the level of the offspring’s DNA. In terms of human laws, marriages between cousins are generally permitted even if children resulting from such marriages will have a Coefficient of Inbreeding of 6.25%.
Inbreeding and the Loss of Genetic Diversity
Once again, for a Coefficient of Inbreeding of 10%, this would mean that for genetic sites that are heterozygous (M/N, carriers) in a common ancestor, there is a 10% chance that these sites will become homozygote (N/N or M/M) in the descendant. In other words, there is on average a 10% net loss of genetic diversity (or gain in genetic uniformity if you will) in the offspring due to matings involving inbreeding. A particular genetic site that has been converted from heterozygous (diversity) to homozygous (uniformity) in an offspring due to inbreeding is said to be identical by descent due to the common ancestor.
Pedigrees and the Coefficient of Inbreeding
 How many generations of a pedigree should be considered when calculating the Coefficient of Inbreeding? The more generations that are available, the more reliable the calculation. Using a small number of generations tends to give coefficient values that are artificially low compared to values obtained when more generations are included. In practical terms, use all the pedigree information that is available. This could be as few as three generation or as many as all the generations in the pedigree going back to the founding animals for the breed.
How many generations of a pedigree should be considered when calculating the Coefficient of Inbreeding? The more generations that are available, the more reliable the calculation. Using a small number of generations tends to give coefficient values that are artificially low compared to values obtained when more generations are included. In practical terms, use all the pedigree information that is available. This could be as few as three generation or as many as all the generations in the pedigree going back to the founding animals for the breed.
Thanks to the power of the internet, many horse pedigrees are available on web sites dedicated to this purpose. Here are several examples:
http://www.allbreedpedigree.com
https://www.aqha.com/-/aqha-records-research
For a given animal, the known pedigree is presented, common ancestors are noted, and often inbreeding coefficients can be calculated for the known generations. A useful function for breeders is the ability to calculate and compare inbreeding coefficients for possible future breedings (“virtual breeding” function).
Using Inbreeding Coefficients for Breeding Decisions
 Inbreeding coefficients, used correctly, are a powerful tool for breeders when it is the time to choose the parents of the future generation of horses. In a general fashion, the inbreeding coefficient is an indication of the global state of genomic health of an animal. The genome is the sum total of all the genes that are necessary for the creation as well as the function of an animal. More specifically (as mentioned above), the inbreeding coefficient represents a numerical percentage, based on the analysis of a pedigree, that estimates the loss of genetic variation in an individual caused by the fact of having common ancestors on both the paternal and the maternal sides of the pedigree. Having common ancestors on the two sides of the pedigree will result in a percentage of genetic sites that were heterozygous (M/N) in the common ancestor to become homozygote (either N/N or M/M) in the descendant. This condition is known as being identical by descent due to the fact of having a common ancestor.
Inbreeding coefficients, used correctly, are a powerful tool for breeders when it is the time to choose the parents of the future generation of horses. In a general fashion, the inbreeding coefficient is an indication of the global state of genomic health of an animal. The genome is the sum total of all the genes that are necessary for the creation as well as the function of an animal. More specifically (as mentioned above), the inbreeding coefficient represents a numerical percentage, based on the analysis of a pedigree, that estimates the loss of genetic variation in an individual caused by the fact of having common ancestors on both the paternal and the maternal sides of the pedigree. Having common ancestors on the two sides of the pedigree will result in a percentage of genetic sites that were heterozygous (M/N) in the common ancestor to become homozygote (either N/N or M/M) in the descendant. This condition is known as being identical by descent due to the fact of having a common ancestor.
From the genetic perspective, a loss of genetic variability is undesirable as it can result in the condition of inbreeding depression. Inbreeding depression has been well documented for many animal and plant species. Also well documented is the effect of outbreeding or crossbreeding, which will increase the genetic variation in the genome of an animal and result in hybrid vigor (heterosis). Hybrid vigor is the genomic flip side of inbreeding depression.
 It is important to keep in mind that the inbreeding coefficient does not represent the genetic variation that will be found at a specific genetic site or gene, but rather is a global estimation of genetic variation for the genome of an animal. If there are no common ancestors between the paternal side and the maternal side of a pedigree, the offspring will have no loss of genetic variation compared to a standard population. The offspring will then have an inbreeding coefficient of 0. If there are common ancestors on both the paternal and the maternal side of the pedigree, there is now the potential of loss of genetic variation (genetic identity by descent), and the potential for inbreeding depression. The Coefficient of Inbreeding is now a positive number greater than 0, often expressed as a percent. It is estimated that for every 1% increase in the inbreeding coefficient there is a 1% reduction in whatever trait is being measured. In practical terms the inbreeding coefficient is most useful for estimating the effects of recent inbreeding.
It is important to keep in mind that the inbreeding coefficient does not represent the genetic variation that will be found at a specific genetic site or gene, but rather is a global estimation of genetic variation for the genome of an animal. If there are no common ancestors between the paternal side and the maternal side of a pedigree, the offspring will have no loss of genetic variation compared to a standard population. The offspring will then have an inbreeding coefficient of 0. If there are common ancestors on both the paternal and the maternal side of the pedigree, there is now the potential of loss of genetic variation (genetic identity by descent), and the potential for inbreeding depression. The Coefficient of Inbreeding is now a positive number greater than 0, often expressed as a percent. It is estimated that for every 1% increase in the inbreeding coefficient there is a 1% reduction in whatever trait is being measured. In practical terms the inbreeding coefficient is most useful for estimating the effects of recent inbreeding.
It is not surprising that Mother Nature likes genetic variation as this is the long-term key to the survival and evolution of a species. Unfortunately for our domestic animals, breed standards favor phenotypic uniformity (breeding for type), and in order to achieve this uniformity, a certain level of inbreeding and reduced genetic variation is involved.
Interpretation of Inbreeding Coefficients
 An elevated inbreeding coefficient for an animal indicates that the undesirable effects of inbreeding (i.e. inbreeding depression) will start to be evident. On the other hand, an elevated inbreeding coefficient will increase the chances that desirable traits associated with the breed in question will be fixed. Thus, the inbreeding coefficient can be viewed as a compromise. The deleterious effects associated with inbreeding start to be seen when the coefficient of inbreeding is higher than 5%, which is just below the value obtained for the offspring of a mating between two cousins (=6.25%).
An elevated inbreeding coefficient for an animal indicates that the undesirable effects of inbreeding (i.e. inbreeding depression) will start to be evident. On the other hand, an elevated inbreeding coefficient will increase the chances that desirable traits associated with the breed in question will be fixed. Thus, the inbreeding coefficient can be viewed as a compromise. The deleterious effects associated with inbreeding start to be seen when the coefficient of inbreeding is higher than 5%, which is just below the value obtained for the offspring of a mating between two cousins (=6.25%).
It is advised to maintain a coefficient of inbreeding that is below 10% which should allow a number of desired traits to be fixed without allowing the undesirable effects of inbreeding to become too pronounced. Incestuous crosses resulting in offspring with coefficients of inbreeding above 12.5% should not be performed; these include parent-offspring, brother-sister, grandparent-grandchild, half-brother-half-sister matings. In practice it is recommended to choose crosses that will result in offspring that have reduced coefficients of inbreeding compared to the average of the breed in question. If a number of breeding possibilities are available that will reduce the average inbreeding coefficient in the offspring compared to the breed average, then ideally the breeding that will result in the lowest inbreeding coefficient while still maintaining the desired traits for the breed is recommended.
 The natural history of the evolution of the horse is particular amongst the domestic animals. The archaeological record shows that the first ancestor of the horse was a small three-toed herbivore that lived in the forests of the Americas about 50 million years ago. This animal is given the poetical name of Eohippus, or “dawn horse”. Eohippus was an ungulate (hooved) animal, meaning that it walked on the tips of its toes which were hooved. Eohippus would give rise to the order Perissodactyles (perisso-, odd numbered; dactyls, toes), the ungulate species that walk on an odd number (one or three) toes per limb. These species include the modern-day tapirs, five species of rhinoceros, and seven species of equids (horses and related animals).
The natural history of the evolution of the horse is particular amongst the domestic animals. The archaeological record shows that the first ancestor of the horse was a small three-toed herbivore that lived in the forests of the Americas about 50 million years ago. This animal is given the poetical name of Eohippus, or “dawn horse”. Eohippus was an ungulate (hooved) animal, meaning that it walked on the tips of its toes which were hooved. Eohippus would give rise to the order Perissodactyles (perisso-, odd numbered; dactyls, toes), the ungulate species that walk on an odd number (one or three) toes per limb. These species include the modern-day tapirs, five species of rhinoceros, and seven species of equids (horses and related animals).